VETENSKAPLIGA KLARGÖRANDEN ANGÅENDEDET ANOMALA FÄRGSEENDET
Jag skall nu skissera de viktigaste av de idéer rörande färgseendets fysikaliska och fysiologiska förutsättningar, som leder fram till en i första approximation giltig teoretisk modell för de vanliga formerna av anomalt färgseende.
SPEKTRUM OCH DESS BETYDELSEIsaak Newtons viktiga bidrag till optiken var hans idé om en enkel, linjär beskrivning av ljusets växelverkan med materia. När ljusstrålning, i form av belysning, träffar de materiella föremålen, och därvid delvis absorberas, delvis passerar igenom och delvis återkastas från deras ytor, så modifieras ljuset. Att vi iakttar skilda färger hos föremålen framför oss, beror på att det ljus de återkastar, och som når våra ögon, modifierats på skilda sätt, alltefter de optiska egenskaperna hos respektive föremåls yta. Newton kom på idén, bestyrkt genom en rad eleganta experimentella demonstrationer, att ljusets mångahanda modifikationer skulle kunna beskrivas med referens till en serie "enkla ljussorter" – vad som sedermera har kommit att kallas monokromatiskt ljus; ljus av bestämd våglängd, eller frekvens. Monokromatiskt ljus kan framställas genom att man låter en smal stråle av vitt ljus (från solen eller en glödlampa) brytas i ett glasprisma, eller reflekteras mot ett finritsat gitter. Därvid uppstår ett spektrum av färger, där varje position – och alltså även varje färg – motsvarar en bestämd våglängd. Vill man nu beskriva hur en viss föremålsyta modifierar infallande ljus, så räcker det att undersöka i vilken grad den återkastar monokromatiskt ljus vid olika våglängder. Ljus i vissa våglängdsområden kanske visar sig absorberas kraftigt av materialet, medan ljus i andra våglängdsområden till största delen återkastas. Reflektionsfaktorn, som funktion av våglängden, karakteriserar ytans optiska egenskaper och är motsvarigheten till vad vi uppfattar som ytans färg. Detta var Newtons centrala idé om vårt färgseendes fysikaliska aspekt, som han formulerade i följande ordalag: "Colours in the Object are nothing but a Disposition to reflect this or that sort of Rays more copiously than the rest; in the Rays they are nothing but their Disposition to propagate this or that Motion into the Sensorium, and in the Sensorium they are Sensations of those Motions under the Forms of Colours." Det är denna modell för ljusets roll vid fägseendet, som ligger till grund för senare tiders exakta forskning och har föranlett att så mycket av denna ägnats åt att utröna hur vårt synorgan reagerar på monokromatiskt ljus. Förhoppningen bakom detta är att det skall vara tillräckligt, dvs har man väl klarlagt effekten av monokromatiskt ljus, så kan man genom en enkel summering beräkna vilken effekt naturligt förekommande ljus har, som ju är långt ifrån monokromatiskt.
Hur ett spektrum ser ut (dvs vilka kulörer man förknippar med monokromatiskt ljus av olika våglängder) har alltid varit ett viktigt test på anomalt färgseende. Likaledes säger den spektrala diskriminationsförmågan (dvs hur mycket man, vid olika våglängder, behöver ändra våglängden för att försökspersonen skall uppfatta en ändring av kulören) en hel del om arten och graden av anomali. Som vi minns beskrev Dalton hur spektrum såg ut för honom, och konstaterade att han bara såg det i olika mättnadsgrader av gult och blått. Hur ser spektrum ut för den normalseende? Det är, noga besett, ett band med kontinuerligt skiftande kulörton, där emellertid vissa toner dominerar. Newton, för sin del, ansåg det berättigat att särskilja sju kulörområden: rött, orange, gult, grönt, blått, indigo, violett. Man kunde nu, som en förklaring till detta utseende, tänka sig att i ögat funnes sju slags receptorer, känsliga för ljus inom respektive våglängdsintervall, svarande mot de antagna "grundfärgerna". Med andra ord, att ögat skulle utföra en grov spektralanalys av det mottagna ljuset. Det spektrum som Newton arbetade med – varvid solljus släpptes in genom ett litet hål i en fönsterlucka och därefter bröts genom ett glasprisma – var inte särskilt spektralrent (se Bouma 1971, p 173). Kanske var det därför som Newton fann anledning att särskilja så många som sju olika kulörer. Betraktar man ett spektrum som helhet, på lite avstånd, är det annars påfallande att det domineras av tre färger: violett, grönt och rött. Mellan det violetta och det gröna finns en smal övergångszon av blåa toner; mellan det gröna och det röda likaledes en smal övergångszon av gula toner (gulgrönt, gult, orange). Dessa övergångszoner är smalare ju renare spektrum är. Den mångkunnige Thomas Young konstaterade, hundra år efter Newtons klarlägganden, att det faktiskt skulle räcka med tre slags receptorer i ögat, för att kulörvariationerna över spektrum skulle kunna beskrivas entydigt. Nämligen om man inte binder sig vid antagandet, att varje kulörton måste ha sin egen receptor, utan i stället tänker sig att de varierande kulörerna svarar mot olika proportioner av excitering av de tre receptortyperna. Detta är den s.k. tre-receptor-teorin, som först ytterligare hundra år senare anammades och konkretiserades av den skotske fysikern och matematikern James Clerk Maxwell och av den tyske medicinaren och fysikern Hermann von Helmholtz. Därmed inleddes det exakta experimentella utforskandet av vårt synsinnes funktionssätt.
TRE-RECEPTOR-TEORINDen modell för färgseendet, som i sina grunddrag skisserades av Young, Maxwell och Helmholtz, är enkel och lättbegriplig. Vi har tre receptortyper i näthinnan, inte för skilda våglängdsband, men med maximum vid skilda våglängder. Ett vid korta våglängder, ett vid mellanvåglängder och ett vid långa våglängder. Kalla dem S-, M- och L-receptorn. Om nu ögat träffas av monokromatiskt långvågigt ljus, så att huvudsakligen bara L-receptorn stimuleras, uppfattar försökspersonen ett mörkt, djupt mättat rött. Stimuleras ögat av kortvågigt ljus, så att huvudsakligen S-receptorn exciteras, ser man ett likaledes mörkt, mättat violett. Vid lämpligt vald våglängd i mellanregistret kan man excitera M-receptorn, varvid man ser grönt (ehuru ljusare och inte fullt lika mättat, eftersom oundvikligen samtidigt även L- och S-receptorerna exciteras). Väljer man i stället en våglängd där L- och M-receptorn exciteras i ungefär samma grad, ser försökspersonen gult. Dominerar excitation av L-receptorn, blir färgen orange (dvs rödgult), överväger i stället excitationen av M-receptorn ser vederbörande gröngult. Träffas retina av ljus med en våglängd, vid vilken inte bara S- utan i någon grad även M-receptorn stimuleras, så ser man blått, eller, vid starkare excitation av M-receptorn, turkos. På så sätt kan kulörvariationen över spektrum förklaras. Förutsättningen är att man accepterar tanken, att förnimmelsen av helt nya kulörer –- exempelvis gult – kan framkallas genom den samtidiga excitationen av olika slags receptorer, och alltså inte behöver ha någon särskild egen receptor i retina. Vi upplever ju inte gult som "röd-grönt"! Observera att det av det skälet måste anses vara en vilseledande popularisering, att kalla de tre receptorslagen för "röd-receptorer, grön-receptorer och blå-receptorer". Vi har inga receptorer (mottagare) för färg, i bemärkelsen kvalitativ förnimmelse. Vi har receptorer för strålning. Belyses retina med vitt ljus, dvs ljusstrålning omfattande hela det synliga våglängdsbandet, exciteras alla tre receptortyperna lika mycket – det är just den stimulering som motsvarar förnimmelsen av vitt, dvs av neutrala (akromatiska) färger. Även "vitt" är en kvalitativt enhetlig färg, inte något slags "röd-grön-blått".
Denna schematiska modell för färgseendet har under årens lopp, genom sofistikerade experimentella undersökningar, alltmer konkretiserats och detaljerats. Förutsättningen har varit avancerad elektronik, som möjliggjort elektrofysiologiska undersökningar med mikroelektroder, varigenom responsen i enstaka nervceller kunnat registreras, på olika nivåer i systemet öga-hjärna. Histologiska studier av ögats näthinna med elektronmikroskop, har avslöjat den fina hexagonala mosaiken av stavar och tappar, under flera lager av genomskinliga nervceller. Stavarna gör tjänst vid skymningsseende och innehåller det ljuskänsliga pigmentet rodopsin, som har sitt absorptionsmaximum vid 505 nm, dvs i det blågröna området av spektrum. Det finns cirka 120 miljoner stavar, fördelade över retina. Tapparna, som aktiveras först vid högre belysningsnivåer och sålunda svarar för dagsljusseendet, är av tre slag, alltefter det fotopigment de innehåller. Enligt mätningar (på apa) av Baylor et al (1987) ligger absorptionsmaxima vid 430 nm, 530 nm resp. 560 nm. Vanligast förekommande är de sistnämnda, alltså L-receptorerna. Därnäst kommer M-receptorerna. S-receptorerna är relativt sparsamt förekommande (mindre än 10%). Tapparna finns huvudsakligen i den centrala delen av näthinnan, den s.k. gula fläcken (macula lutea). I mitten av denna, fovea centralis, är tapparna särskilt små och sitter mycket tätt, uppskattningsvis 147000 per kvadratmillimeter. Totalt, över retina, finns 4-7 miljoner tappar. I mitten av fovea, motsvarande en synvinkel på en grad, har vi vårt skarpaste seende. När vi vill fixera ett föremål riktar vi spontant blicken så att den optiska bilden av föremålet hamnar på fovea. Det perifera seendet är visserligen mer oskarpt, men i gengäld ytterst vidvinkligt och bland annat med stor förmåga att uppfatta ting som rör sig. Vid det svaga skymningsljuset, då stavarna tar över uppgiften att detektera ljusstrålning, har man sin största känslighet perifert. Man kan exempelvis uppfatta stjärnor, vars sken är så svagt att man inte ser dem om man fixerar dem. Eftersom det tycks som om åtminstone vårt foveala färgseende baseras på förekomsten av ljuskänsliga receptorer av tre skilda slag kallas det "trikromatiskt". Å andra sidan är förmågan att vid perifert seende uppfatta och korrekt identifiera färger avsevärt mindre än vid centralt, eftersom de perifera delarna av näthinnan är så relativt glest befolkade av tappar. Där krävs dessutom, av samma skäl, avsevärt större area hos den fläck, vars färg man skall varsebli. Färgerna upplevs mindre mättade. I synnerhet gäller detta röda och gröna, som ser mer gula ut, medan violett ter sig blått. Vi har med andra ord en tendens till röd-grön-anomali ut mot perifierien av synfältet. Att man koncentrerat så mycket av psykofysiska mätningar till centralt seende, beror på att man där har i olika avseenden säkrast och mest väldefinierat seende. Våglängds-diskriminationen är optimal för en synvinkel på mellan 1 och 3 grader. Därunder, och däröver, minskar den – dvs det krävs större förändringar av våglängden hos en monokromatisk stimulus, för att försökspersonen skall uppfatta en skillnad. Även luminansen har betydelse för att vi skall uppfatta kulör hos mycket små fält (exempelvis avlägsna ljussignaler). Minst luminans för att uppfattas kräver rött. För grönt och blått ljus ligger tröskeln för synlighet högre. Vid för låg luminans uppfattas det gröna som vitt, men det gäller faktiskt även vid för hög luminans. Så över en viss nivå gagnar det inte att öka styrkan på en grön signal, för att säkerställa att den skall uppfattas som grön och icke som vit. Stjärnor, vilkas strålning har maximum i det gröna området av spektrum uppfattas likväl inte som "gröna" utan som vita. Nå, det var en liten utvikning om det normala färgseendets fysiologi. Nu åter till de grundläggande aspekterna på den trikromatiska modellen för färgseendet.
KOMPLEMENTARITETAv vad jag sa om att en likformig stimulering, som i samma mån exciterar alla tre slagen av receptorer, uppfattas som vitt, dvs okulört färg, följer en mycket viktig princip för färgseendet (som redan Newton utifrån sin teori insåg) nämligen komplementaritet mellan färgerna. Säg att vi har vitt ljus, dvs ett kontinuerligt någotsånär likafördelat spektrum, omfattande alla våglängder över den för seendet betydelsefulla oktaven 380 - 760 nm. Om vi nu delar upp detta "fullspektrum" i två delar, genom ett snitt vid 500 nm, så kommer den kortvågiga delen att excitera S-receptorerna, alltså uppfattas som violett ljus; resten exciterar M- och L-receptorerna och uppfattas alltså som gult ljus. Med andra ord: Eftersom excitering av alla tre receptorerna innebär vitt, så kommer den färg som svarar mot en simultan excitering av två av receptorerna att vara komplementär till den färg som svarar mot excitation av den tredje. Sålunda är rött och cyan (blågrönt) komplementära. Som grönt uppfattar vi ett ljusflöde som huvudsakligen exciterar M-receptorn. Det är komplementärt till ljus som simultant exciterar S- och L-receptorerna, motsvarande en blåröd kulör, ofta kallad magenta. Man inser att denna komplementaritetsprincip inte bara gäller det idealiserade fallet, utan även det allmänna fallet, då de tre receptorerna exciteras i varierande proportioner. För varje proportion av stimulering finns det en annan proportion som är dess "komplement" i den meningen att de tillsammans motsvarar neutralpunkten, dvs den proportion av exicitering som "vitt ljus" medför. Tre-receptor-teorin ger oss sålunda ett färgsystem, baserat på sex grundfärger, vilka är parvis komplementära: gult och violett, magenta och grönt, cyan och rött. Observera att magenta är en kulör som inte återfinns i spektrum. Det finns ingen våglängd som ensam kan ge en likvärdig stimulering av L- och S-receptorerna. Det är en viktig egenskap hos tre-receptor-teorin, att den innebär att det existerar kulörer, som inte är representerade i spektrum, utan som kräver stimulering med sådant ljus, som innehåller komponenter från skilda våglängdsområden. För att klarlägga färgseendet räcker det med andra ord icke att arbeta enbart med strålar av monokromatiskt ljus.
METAMERIDet tilltalande med tre-receptor-teorin är att den innebär en kraftig begränsning av den mängd spektrala sammansättningar som man behöver studera för att fastlägga färgseendet. Detta på grund av att flera olika spektralsammansättningar av ljuset exciterar de tre slagen av receptorer i exakt samma proportion, dvs under i övrigt lika förhållanden uppväcker samma färgförnimmelse. En konsekvens härav är att två olika färgmaterial, betraktade i en viss belysning, kan se precis likadana ut, dvs ha samma kulör, fastän de i sin modifierande verkan på ljuset är helt olika. (Vilket kan avslöjas, genom att man betraktar dem i annan belysning.) De är "förväxlingsfärger" för den normalseende. Fenomenet kallas "metameri". En varelse utrustad med fyra skilda slag av receptorer, skulle inte tycka att de två nämnda färgmaterialen var lika och förvåna sig över hur "färgblinda" vi människor är. Exempelvis skulle denna varelse inte tycka att den kulör som framställs genom additiv blandning av rött och grönt på en TV-skärm skulle vara en acceptabel imitation av den kulör i naturen som kallas "gult". Men det tycker vi! Man kan säga att det normala färgseendet karakteriseras av sina förväxlingsfärger, på samma sätt som det anomala seendet karakteriseras av sina. Att bestämma vilka optiska stimuli som är kromatiskt ekvivalenta, är ett sätt att kartlägga en individs färgseende. Lägg märke till att detta kan göras utan att man behöver gå in på frågan vad han "egentligen" ser, dvs undersöka vilka färgförnimmelser han förknippar dessa stimuli med. Ty själva experimentet består bara i att fastställa om två intilliggande fält har samma kulör eller inte. Däremot behöver försökspersonen inte yttra sig om vilken kulör fälten har, när de är lika.
DIKROMASI OCH ANOMAL TRIKROMASIVi är nu mogna för att uppställa en schematisk teori för det anomala färgseendet. De tre receptorerna kännetecknas av sin respektive spektrala känslighet. Om nu känslighetskurvan för en av receptortyperna förskjuts i spektrum, så kommer färgseendet att ändras på så sätt att nya par av optiska stimuli blir "förväxlingsfärger". (Det har föreslagit även andra modeller, men detta är den sk. "single-pigment shift model", se Pokorny och Smith 1977) En person med en sådan anomali, skulle alltså tycka att vissa färgprover, som den normalseende påstår är lika, har olika kulör, samtidigt som vissa andra färgprover, vilka för honom ser likadana ut, av den normalseende påstås vara olika. Mycket riktigt visar sig detta svara mot hur en grupp av personer med anomalt färgseende uppför sig. Dessa personer kallas "anomala trikromater". Alltefter om det är L-receptorns känslighetskurva som har förskjutits i riktning mot M-receptorns, eller om det i stället är M-receptorns känslighet som förskjutits i riktning mot L-receptorns, talar man om protanomal trikromasi respektive deuteranomal trikromasi. Om nu förskjutningen går så långt att L- och M-receptorernas känslighetskurvor sammanfaller (dvs om de innehåller samma fotokänsliga pigment) då kommer variationsmöjligheterna i det sätt på vilket receptorsystemet kan exciteras att bli mindre, eftersom L- och M-receptorerna då alltid exciteras i samma grad, oavsett våglängdssammansättningen hos det ljus som belyser dem. För en dikromat finns sålunda optiska stimuli som är omöjliga att skilja, som en trikromat lätt ser skillnad på. Det är just vad som kännetecknar utpräglad anomali. Även dikromaterna kan vara av två slag, alltefter om L+M-receptorernas känslighet motsvarar den normalseendes M-receptorer, eller L-receptorer. I det förra fallet talar man om protanoper, i det senare om deuteranoper. Protanopen kännetecknas av att djupt röda färger blir mycket mörka, eftersom han har ringa känslighet vid långa våglängder. Det föranleder hans svårigheter, när det gäller att uppfatta röda ljussignaler. Utöver de anomala trikromaterna (vilket är en lindrig form av anomali) och dikromaterna, räknas i praktiken som en särskild grupp de s.k. extremt anomala, som visserligen har något skilda känsligheter för L- och M-receptorerna, ehuru denna skillnad är så liten att symptomen blir likartade dikromaternas. Fördelningen, i vår västerländska civilisation, mellan de sex vanliga slagen av medfödd anomali bland män, framgår av följande tabell:
Att anomalt färgseende är så mycket vanligare bland män än bland kvinnor beror på att den gen som bestämmer L- och M-pigmenten finns på X-kromosomen. Kvinnor har två sådana, en från vardera föräldern, och kan sålunda med stor sannolikhet ha en "normal" information beträffande dessa fotopigment. Hos en man som från modern ärver en "anomal" gen blir emellertid denna utslagsgivande. Det är därför som anomalt färgseende ärvs från morfar till dottersöner. En och annan torde väl undra om inte också S-receptorerna kan ha en avvikande känslighetskurva, vilket skulle ge diverse ytterligare typer av anomali. Eller att de skulle helt saknas, vilket skulle medföra "tritanopi", dvs en individ med endast L- och M-receptorer. Denna form av anomali förekommer faktiskt, men är inte tillnärmelsevis lika vanlig som de ovannämnda formerna, beroende på att den gen som bestämmer vilket ljuskänsligt pigment S-receptorn får, inte befinner sig på X-kromosomen utan på kromosom 7 (se Piantanida 1991) och sålunda finns i dubbel uppsättning hos både män och kvinnor. Även total färgblindhet, akromatopsi, förekommer men är mycket sällsynt. Den består då vanligtvis inte i "monokromasi", dvs att vederbörande bara har ett slag av tappar i retina, utan i stället i att personen ifråga endast har skotopiskt seende, dvs endast stavar, och därmed stora svårigheter att överhuvudtaget se på dagsljusnivåer.
NEUTRALPUNKTEN I SPEKTRUMJag beskrev förut hur spektrum ser ut för en normalseende. Hur ser då spektrum ut för en individ med utpräglat anomalt färgseende (en protanop eller deuteranop) ? Man kunde naivt tro att den som saknar normala L-receptorer skulle vara "röd-blind" och se världen i varierande nyanser av grönt, blått och violett. Likaledes, att den som saknar normala M-receptorer skulle vara "grön-blind" och se världen i rött och violett. Men den slutsatsen är ohållbar, av ett mycket enkelt skäl. Nämligen att vitt ljus uppfattas som vitt. Med andra ord: lika stimulering av de två receptortyperna motsvarar okulörthet. S-receptorn vet vi motsvarar violett; då måste den andra receptortypen hos dikromaten motsvara komplementfärgen till violett, nämligen gult. Med hänvisning till den enkla modellen ovan, kan man säga, att om båda receptorsystemen (L- och M- ) finns hos dikromaten, men har kommit att innehålla ett och samma fotopigment, så betyder ju det att de alltid exciteras i samma grad, vilket enligt modellen skulle innebära förnimmelsen av gult. Enligt det resonemanget skulle dikromaten, såväl protanopen som deuteranopen, vara oförmögen att se vare sig rött eller grönt, eftersom det inte finns något ljus som kan excitera L- receptorerna i hög grad och samtidigt M-receptorerna i ringa grad, eller vice versa. Vid båda fallen av anomali ser vederbörande alltså spektrum endast i nyanser av gult och blått, så som John Dalton på sin tid beskrev det. Det intressantaste draget i dikromatens uppfattning av spektrum är emellertid att det finns en vitpunkt, en viss välbestämd våglängd, som för honom är helt neutral, dvs monokromatiskt ljus av denna enda våglängd ger samma färgintryck som ljus sammansatt av alla våglängder! Bestämning av en eventuell vitpunkt i spektrum anses vara en säker metod att fastställa dikromasi. Dess läge skiljer sig något för protanoper och deuteranoper. Exakt var den ligger beror på vilken vitstandard man valt som referens och låter försökspersonerna adaptera till. Om vitreferensen är 6500 K temperaturstrålning, ligger vitpunkten genomsnittligt för protanoper vid 492 nm och för deuteranomper vid 498 nm, se Ruddock 1991, sid 12. (I båda fallen förskjuts vitpunkten mot längre våglängder, om man har varmare ljus.) I purpur finns en motsvarande vitpunkt. Det finns m.a.o. rosa toner som den anomale uppfattar som vitt.
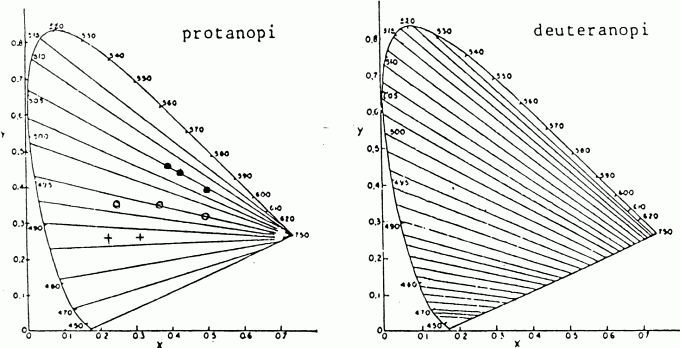
FÖRVÄXLINGSLINJER I KROMATICITETSDIAGRAMMETDen trikromatiska principen innebär bl.a. att mångfalden av särskiljbara färgstimuli (ljusstrålning med varierande spektral energifördelning) kan representeras i ett s.k. kromaticitetsdiagram. Varje punkt i diagrammet anger de stimuli som (efter lämplig justering av ljusheten) är "förväxlingsfärger" för en normalseende. En dikromat förväxlar emellertid även vissa av de stimuli som en normalseende ser skillnad på och det visar sig att dessa "förväxlingsfärger" ligger på i stort sett räta linjer genom kromaticitetsdiagrammet. (s k confusion lines). Det överensstämmer med vad man skulle förvänta sig, om dikromasien består i att ett av de tre primära receptorsystemen saknas. Med andra ord, att dikromasin skulle vara att anse som en reduktionsform av trikromasin (Scheibner). Förväxlingslinjerna löper något annorlunda för protanoper och deuteranoper (se figur.)
Som synes löper vissa förväxlingslinjer från rött, via orange och gult upp till gulgrönt och det även vid avsevärd mättnad. Till och med monokromatiskt ljus ser, vid våglängder > 540 nm, i kulörhänseende exakt lika ut. Någonstans mitt i diagrammet ligger vitpunkten. Den anger kromaticiteten för de stimuli som uppfattas som neutrala, okulörta. Den förväxlingslinje som går genom vitpunkten anger stimuli som för oss är kulörta (och komplementära), men för dikromaten är neutrala. Med andra ord (såsom framgår av figuren, öppna ringar) en röd och en blågrön kulör. Som synes är den extremt röda delen av spektrum ( > 650 nm) för en protanop komplementär (med avseende på ljuskälla E) till ljus av våglängden 494 nm. Den våglängden i spektrum är alltså neutralvit. Detta är, som sagt, ett av kännetecknen på en dikromat: att han har en vitpunkt i spektrum, i det område där en trikromat ser en blågrön kulör. Vitpunkten är dessutom relativt välbestämd, ty den anomale har sin största spektrala diskriminationsförmåga i just det området. De anomala trikromaterna har i viss utsträckning samma förväxlingsfärger som dikromaterna. De har ingen vitpunkt i spektrum, men blågröna färger är kulörsvaga. Omättade färger kan lätt förväxlas utefter samma riktningar i kromaticitetsdiagrammet som de vilka gäller för dikromater. Det är utifrån den principen som s.k. PIC-tavlor ("pseudo-isochromatic charts") konstruerats. Små färgprickar i olika nyanser bildar vissa figurer (vanligtvis bokstäver eller siffror) som den anomale icke ser, eftersom de framträder mot bakgrund av prickar i förväxlingsfärgen. Till yttermera visso framträder vissa förvillande figurer, beroende på varierande ljushet hos prickarna. PIC-tavlor är användbara för att påvisa såväl anomal trikromasi som dikromasi. Det s.k. anomaloskopet bygger på samma princip. I det fallet presenteras ett lysande fält, som består av två halvor. På den ena presenteras ljus av våglängden 589 nm (gula natriumlinjen) , på den andra en variabel blandning av 546 nm och 671 nm. Det gäller att ställa in blandningen så att de två halvorna smälter samman till ett enda gult fält (s.k. "Rayleigh-match"). En dikromat accepterar vilken som helst blandning. En anomal trikromat har en bestämd inställning, som emellertid avviker från de normalseendes. Även datorskärmar (CRT-monitorer) har använts för att fastställa förväxlingsfärger. Så exempelvis i en studie av Derefeldt et al. En lovande metod att direkt bestämma hela förväxlingslinjer (i stället för enstaka färgpar) har utarbetats vid Biofysikgruppen vid Oslo universitet.
DE ANOMALAS DISKRIMINATIONSFÖRMÅGAFörmågan att uppfatta små ändringar i våglängd som en kulörändring är för en normalseende i vissa områden av spektrum mycket stor. Under optimala försöksbetingelser kan det röra sig om så litet som 1 nm i det blågröna och i det gula området. Ett kännetecken för de färganomala är att diskriminationsförmågan är avsevärt sämre, i synnerhet i det gula och gulröda området. För protanoper och deuteranoper är den spektrala diskriminationsförmågan mycket ringa. Det är bara i området kring neutralpunkten som känsligheten kan vara i bästa fall nedåt 4 nm (dvs jämförbar med vad en normalseende har över en stor del av spektrum). Även anomala trikromater har genomgående sämre diskriminationsförmåga än normala trikromater (det är ett av skälen till att deras anomali ofta undgått deras uppmärksamhet i praktiska livet. De märker inte skillnaden mellan sina egna färgbedömningar och andras.) Framför allt varierar den betydligt kraftigare än normalt med våglängden. Av jämförbar storleksordning med det normala är den bara i området kring 500 nm. En viss diskriminationsförmåga finns i det gulröda området (kring 600 nm), men i det violetta, det gulgröna och det röda området av spektrum fordras mycket stora våglängdsändringar för att det skall varseblis som en kulörförändring. Ett färgblindhetstest som bygger på den nedsatta diskriminationsförmågan hos anomala är det s.k. "Farnsworth 100 hue test" – där uppgiften är att ordna en uppsättning små färgprover, med tämligen omättade kulörer, till en kulörtonkrets.
OPPONENTFÄRGTEORIMöjligheten att beskriva spektrums utseende i termer av binärkulörer, dvs som rödgult, gulgrönt, grönblått, blårött, har inspirerat en modell för färgseendet, som bygger på opponent-processer. Eftersom några röd-gröna kulörer inte finns, utan rött och grönt avlöser varandra, när man vandrar genom spektrums våglängder, så kan man tänka sig en nevronal process (rg-processen) som ger positiv respons för rött (exempelvis i form av ökad nervaktivitet) och negativ respons för grönt (i form av minskad aktivitet, i förhållande till en neutralnivå). På samma sätt definieras en yb-process. Den rent gula kulören, som är varken rödgul eller gröngul, eller en rent blå kulör, som är varken grönblå eller rödblå, förnimmes om rg-processen ligger på neutralnivån. På samma sätt förnimmes en rent grön eller rent röd kulör, när yb-processen ligger på neutralnivån. Om både rg- och yb-processen ligger på neutralnivån, blir synintrycket okulört (akromatiskt). Det är fallet vid belysning av receptorerna med vitt ljus. Denna teori lanserades av Ewald Hering, som såg en generell princip i att ett organiskt system, som befinner sig i jämvikt med omgivningen, reagerar på en förändring i det yttre kraftfältet (i detta fall strålningen som träffar näthinnan) med ökad eller minskad metabolism för att återupprätta jämvikten. Herings opponentfärg-teori räknar sålunda med fyra "elementarkulörer" i stället för tre (eller sex) som det tidigare beskrivna trikromatiska systemet. Hur går det ihop? Alltför mycket talar för att den primära receptionen av strålning på näthinnan sker i tre typer av receptorer. Men man kan tänka sig att i de efterföljande lagren av nervvävnad sker ett slags "signalbear-betning" där de initiala responserna omvandlas till opponenta responser. Man har funnit elektrofysiologiska belägg för att det faktiskt fungerar så. Denna "två-stegs-modell" har blivit den allmänt vedertagna. (Nyligen har forskarparet De Valois lanserat en tre-stegs-modell.) Eftersom de fyra elementarkulörerna Y,B,R,G, är opponenta innebär det egentligen bara två dimensioner: den röd-gröna respektive den blå-gula. Den tredje dimension, som erfordras för att representera mångfalden av möjliga nyanser, ombesörjs i Herings teori av en sv-process, som motsvarar dimensionen svart/vitt, dvs gråskalan. Man kunde tänka sig att anomalt färgseende kunde ha sin grund i att rg-processen var defekt, snarare än att det skulle handla om brist på ett fotopigment i de primära receptorerna. Om rg-processen alltid står i neutralläget, innebär det ju att endast kulörer i dimensionen blått/gult förnimms. Mycket talar dock för att de sedvanliga nedärvda formerna av anomali väsentligen har med just fotopigmenten att göra. (Se vidare diskussionen hos Pokorny och Smith, 1977)
ANGÅENDE RÖDHETEN I DET VIOLETTAVarför den kortvågiga delen av spektrum av normalseende uppfattas i en kulör som vi kan beskriva som rödaktigt blå, är en fråga som inte lösts, fysiologiskt. Det finns tre alternativa förklaringar: Antingen har L-receptorns pigment ett svagt absorptionsmaximum även vid korta våglängder, eller så innehåller L-receptorn en ringa inblandning av S-receptorns pigment. Alternativt sker en koppling på nevronal nivå, så att S-receptorerna i någon grad kan inverka positivt stimulerande på rg-processen. Vilketdera som är fallet, vet man inte. Det kan finnas andra förklaringar. Exempelvis, att den förnimmelse som excitation av enbart S-receptorerna ger upphov till är just denna kulör vi kallar "violett", dvs en rödblå kulör. Mot den sistnämnda hypotesen talar emellertid det faktum, att de flesta anomala inte ser denna rödhet i det kortvågiga området. De kallar inte kortvågigt ljus för rödblått. De ser inte skillnad på ett syrenlila färgprov och ett blått. Dessutom är det ett faktum, att normalseende har denna lilla defekt i foveolan, dvs vid mycket små synvinklar i synriktningen. Lägger man upp stenkulor i olika färger, inklusive en violett och en ljusblå kula och sedan betraktar dem på en tjugo, trettio meters håll, så ser man inte längre den violetta kulan som violett, utan som blå!
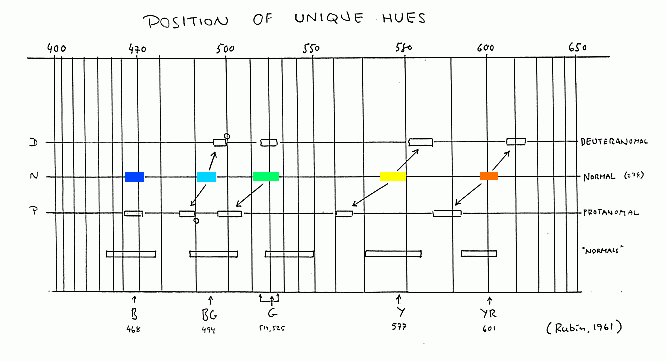
ELEMENTARKULÖRERNAS POSITIONER I SPEKTRUMOpponentfärgteorin gör det intressant att ta reda på vid vilka våglängder som noll-passagerna i rg-processen resp. yb-processen inträffar. Med andra ord ta reda på var en försöksperson upplever att elementarkulörerna (de s.k. "unique hues") befinner sig i spektrum. Den statistiskt mest omfattande undersökningen av den saken gjordes av M.L.Rubin (1961). Den baserades på 278 normalseende, 12 protanomala och 32 deuteranomala försökspersoner. Rubin fann tämligen välbestämda positioner för "unique hues" (Y,G,B) och även för "binary balanced hues" (YR,BG) - dvs den kulör som är lika röd som gul, respektive lika grön som den är blå. Den röda elementarkulören går däremot inte att hitta en bestämd position för, eftersom kulören vid höga våglängder blir djupröd, men aldrig går över i ett definitivt blårött. (Vid låga luminanser, eller om man tidigare uppehållit sig i det gröna området av spektrum, kan man tycka att exempelvis 650nm är rent röd, men vid högre luminanser uppfattas även de längsta våglängderna som aningen gulaktiga, "varmröda".)
Det intressanta var att Rubin fann en klar tendens till bestämda förskjutningar av dessa lägen för anomala trikromater, som framgår av figuren nedan. (Deuteranomala har dock inte möjlighet att fastställa ett bestämt läge för B, eftersom den inte ser den rödhet som kommer in vid låga våglängder och följaktligen inte kan tillämpa kritieriet "varken rödaktig eller grönaktig blå". Dikromater har inte möjlighet att bestämma några positioner för "unique hues". För dem är det neutralpunkten som är det karakteristiska.) Tendensen är emellertid, som synes, att Y och YR återfinns vid lägre våglängder för protanomala, och högre våglängder för deuteranomala, jämfört med den position de har för normalseende. Förskjutningarna är emellertid inte stora nog, i förhållande till interindividuella variationer, för att man skall kunna ställa någon säker diagnos med hjälp av detta slags bestämning. Men det är ett värdefullt komplement till de undersökningar som baseras på förväxlingsfärger. En svårighet ligger också däri, att man måste vara noga med själva proceduren – flera minuters adaptation till mörker och/eller belysning av bestämd kromaticitet, därefter exponering av det lilla ljusfält vars kulör skall bedömas under bestämd kort tid etc. Det gör också en viss skillnad om försökspersonen själv får ställa in monokromatorn och pendlande söka sig fram exempelvis till läget för den våglängd vars gula kulör varken är grönaktigt eller rödaktigt gul. Eller om han presenteras en given kvasi-slumpmässig serie av våglängder i närheten av den unika gula och för var och en får säga om den förefaller grönaktig eller rödaktig, varefter gränsläget bestämmes statistiskt. Men oavsett hur rigorös procedur man använder är det svårt att få ett stabilt resultat vid denna typ av bedömningar (till skillnad från det traditionella matchningarna, exempelvis i Nagel-anomaloskop). Det framgår med önskvärd tydlighet av hur varierande resultaten blivit i den mångfald av undersökningar som gjorts under senare år. (Se översikt i Ayama et al.1987) Laxar et al. (1988) fann dessutom att läget för elementargult och elementarblått i spektrum varierade, även för en och samma försöksperson, under loppet av ett år. Schefrin och Werner (1990) undersökte hur läget för "unique hues" beror av försökspersonens ålder och fann en tydlig effekt för grönt, varvid en lägre våglängd prefereras vid ökande ålder. (Dvs en ljusfläck, med en våglängd som en ung person skulle utvälja som "rent grön", skulle en äldre person kalla "gulgrön".) Vidare finns redan från 1930-talet undersökningar som pekar på två distinkt skilda lägen för den gröna elementarkulören. Den bimodala karaktären hos den statistiska fördelningen av läget för G verifierades i Rubins undersökning, och senare även av Richards (1967). Oavsett ålder finns alltså en avsevärd grupp av normalseende som skulle välja som rent grön en kulör som övriga (merparten) skulle kalla gulaktigt grön. Mina egna erfarenheter av en tentativ studie med en monokromator (och med 2° lysande fläck i mörkt omfält) är, att man genom att få göra upprepade inställningar "kommer fram till" ett läge som känns som det "sanna" och som är det som man vanligtvis direkt ställer in, redan vid första försöket utan speciell omsorg med tid för adaptation m.m. Den inställningen har (för min egen del) varit densamma under ett års förlopp. Intressant är att om man ber försökspersonen ställa in "den bästa representanten för grönt" så väljer de allra flesta oöverlagt en tydligt gulgrön färg, dvs gulare än den kulör de väljer om man uttryckligen tillhåller dem att ställa in den våglängd som har en grön kulör som är varken gulaktigt eller blåaktigt grön. Intressant är också (vilket även Rubin påpekade) att de binära balansfärgerna (YR och BG) går att ställa in med samma reproducerbarhet som elementarkulörerna, till trots av att kriteriet "lika gul som röd" (resp. "lika blå som grön") känns mycket mera vagt, mycket mera som ett godtycke. För de flesta är f.ö. "orange" en rödare kulör än balanskulören YR. Att det skulle finnas betämda positioner i spektrum för Y B och G är en hypotes, som följer av opponent-respons-modellen. Många försökspersoner säger dock spontant att de uppfattar det som två skilda iakttagelser: att se exempelvis var det gröna försvinner (eller börjar) i det gula, respektive var det röda försvinner (eller börjar) i det gula. Det känns svårt att lägga märke till båda sakerna på en och samma gång. Det kan exempelvis förhålla sig så att det finns ett visst intervall som är rent gult, men också (värre) så att det gröna inslaget kommer redan innan det röda har försvunnit. Det finns uppenbarligen en svårkontrollerbar labilitet eller ofrånkomlig obestämdhet i sambandet mellan ljusvåglängd och kulör.
ANGÅENDE KONTRASTKÄNSLIGHETENW.A.Nagel, en av pionjärerna när det gäller utvecklandet av färgsinnestest, var en av de första som drog uppmärksamheten till att färganomala tycks ha förhöjd kontrastkänlighet (Nagel 1904). Han lade märke till att när hans deuteranomala försökspersoner fick betrakta ett rött och ett gult fält, sida vid sida, kallade de alltid det gula för grönt. Men om samma gula prov fick betraktas ensamt, kallade de det gult. Nagel konstruerade ett testkort med bruna prickar mellan röda, i tanken att anomala trikromater spontant skulle ange dem som gröna. I synnerhet har detta visat sig vara fallet vid ljussignaler. En gul lanterna bredvid röd, uppfattas lätt som grön av den anomala. Chapanis (1949) fann, att när två grå prover, med endast liten ljushetsskillnad, presenterades kunde det hända att dikromater och även en del anomala trikromater beskrev det mörkare som "kulört"; och den effekten kunde kastas om, ifall det mörkare provet gjordes ljusare. Egil Hansen, vid Rikshospitalet i Oslo, har arbetat med s.k. florkontrast, som ett supplementärt test vid kliniska undersökningar. (Han bygger därvid vidare på en testmetod, som redan Jennings omnämnt 1896, se Fletcher & Voke 1985, p284) Florkontrast uppstår när exempelvis en grå och en färgad ytor betraktas genom silkespapper eller ett flor, så att färgerna blir mycket omättade (Hansen 1976). Simultankontrast (eller färginduktion, som det stundom också kallas) är ett välkänt fenomen, vad gäller normalt färgseende, men de nämnda forskarna vill göra gällande att det är speciellt utpräglat hos färganomala. Huruvida det verkligen förhåller sig så är emellertid omdiskuterat. Jo Ann Kinney fann i en undersökning (1967) att blått är den kulör som effektivast inducerar kulör i ett testfält, omgivet av det blå. Det gäller för normalseende i samma mån som för den deuteranop hon noga undersökte. När testfältet hade en kromaticitet motsvarande dagsljus beskrev försökspersonen den inducerade kulören som tämligen omättad gul. Intressant nog var det emellertid inte möjligt att omvänt, med ett gult, orange eller rött omfält, inducera en blå ton i testfältet. Min egen erfarenhet är att somliga försökspersoner kan göra intryck av förhöjd kontrastkänslighet, men att det troligtvis kan förklaras av att de är så uppmärksamma på och beredda att ta fasta på minsta lilla kulörnyans. De kallar ogärna ett grått färgprov för grått – tycker sig uppfatta en kulörton hos det. När jag tillsammans med en försökperson genom fönstret betraktade taken på husen mitt emot, på andra sidan gatan – det var ett svart, ett rödbrunt och mellan dem ett grått plåttak – betecknade han med bestämdhet det mellanliggande taket som grönt. Det stod han för, var inte villig att ändra sig, när jag påpekade att det i mina ögon var grått, svagt brunaktigt grått. Med ett vitt omfält kan man inducera in mörker (eller svarthet) i ett gult infält. Det antar då en grönaktig ton. På samma sätt kan ett lysande, klart orange, förvandlas till brunt. De försökspersoner jag visat detta fenomen för har spontant rapporterat att de såg grönheten, respektive förvandligen till brunt. (En försöksperson sade att det gula blev grönaktigt "på ett konstigt sätt".)
ANGÅENDE BEROENDET AV FÄRGFÄLTETS STORLEKSom jag inledningsvis nämnde säger sig de flesta försökspersoner ha svårt att bestämma färgen hos små ytor och har behov av att få studera dem på närmare håll för att kunna uttala sig om färgen. En försöksperson konstaterar: "Ser jag alla färglapparna i en hög på avstånd, då ser jag inte de gröna. Men får jag betrakta lapparna på nära håll, helst en och en, då ser jag de gröna, och när jag väl fått syn på dem, då ser jag dem även på längre håll." Detta beroende av storleken hos det färgfält vars färg skall bestämmas är ett intressant fenomen (Ruddock 1991,13f). Nagel (1905) fann, när han utarbetade sitt anomaloskop, att han själv var deuteranop vid 2° storlek på färgfältet, men var deuteranomal vid 10° synvinkel. Vid undersökning fann hann att detsamma gällde 30 andra dikromater. Bland dessa fann han ingen som misslyckades med att identifiera diverse schatteringar av rött, när en tillräckligt stor yta av retina stimulerades. Smith och Pokorny (1977) gjorde Rayleigh-matcher vid en serie storlekar på färgfältet, i ett experiment med sju protanoper och elva deuteranoper och fann att när synvinkeln ökades från 1° till 8° blev försökspersonernas matchningar trikromatiska. Det har framkastats att denna trikromasi vid större synvinkel skulle ha att göra med att stavar då kunde spela rollen av "tredje receptortyp". Det har emellertid visats av Nagy (1980) att tendensen till trikromasi kvarstår även när stavarna är försatta ur spel genom blekning. Nagy konstaterar även att om de två stimuli, som skall matchas (den gula respektive den av rött och grönt sammansatta), presenteras alternerande på samma fläck, i stället för som två intilliggande fält, blir noggrannheten i matchen bättre. Breton och Cowan (1981) fann, i sin undersökning av Rayleigh-matcher vid varierande luminansnivåer, belägg för att stavarna medverkar vid låga nivåer. Vid högre luminansnivåer blir inställningarna för match emellertid annorlunda, vilket indikerar att därvid trots allt ett tredje pigment finns i några av tapparna. Exempelvis det pigment som kännetecknar en anomala trikromat. Merparten av dem som klassats som dikromater skulle alltså betecknas som extremt anomala trikromater. Montag (1994) konstaterar detsamma, och tillägger att det inte bara är vid små stimuli som de förmenta dikromaterna har svårigheter att genomföra en tredimensionell kategorisering, utan även vid kort presentationstid (60 ms). De nämnda resultaten är av vikt, ty all diagnos av anomalt färgseende bygger på användandet av anomaloskop, med matchning av intilliggande fält och 2° synvinkel. De tidigare angivna procentuella andelarna av dikromater bland de anomala hänför sig sålunda till små stimuli. I och med att en stor andel av dem som klassificerats som dikromater är kapabla att särskilja rött och grönt vid större stimuli, finns ingen anledning förmoda att de inte skulle vara bekanta med förnimmelserna "rött" och "grönt", dvs att de menar detsamma med dessa ord, som vad de normalseende gör. Jag återkommer till den saken i den avslutande diskussionen.
INDIVIDUELLA VARIATIONER I FÄRGSEENDETSom jag nämnde i introduktionen uppvisar färgseendet, redan inom ramen för vad som diagnosticeras som "normalt" stora individuella variationer. Vissa data pekar på att det eventuellt skulle finnas två klart åtskilda typer av normalseende (Richards 1967). De skiljer sig från varandra bl.a. däri att de har olika uppfattning om var den rent gröna elementarkulören återfinns i spektrum. (Se ovan!) Michael Webster och Donald McLeod redovisade i en artikel 1988 en faktoranalytisk studie av mätdata för de 49 observatörer som ligger till grund för CIE:s 10° standardobservatör. De fick fram ett tiotal faktorer som kan vara teoretiskt intressanta. Sex av dessa kunde identifieras som variation i gula fläckens pigmenttäthet, linsens färg, spektrala positionen hos respektive fotopigment samt medverkan av stavarna i färgseendet. Denna undersökning gällde individer som räknas som normalseende, men de nämnda faktorerna medför likaväl individuella variationer mellan personer med anomal färgseende. De Valois (1993) nämner att vissa resultat tyder på att det finns två "normala" L-receptor-pigment, med några få nanometers skillnad i absorptionsmaximums läge. Eventuellt även två M-receptor-pigment. Intressant i detta sammanhang är även den undersökning som Danny Rich och Joelyn Jalijali redovisar (1995) som avsåg matchning av neutralpunkten på data-skärmar. En serie vita och grå prover, i en belysning som hade en färgtemperatur på 9000K, jämfördes med en rgb-blandning på skärmen. Resultatet uppvisade stor spridning normalseende emellan. En anomal försöksperson avvek inte påfallande från den gemensamma fördelningen. Dessa faktorer som medför individuella variationer det "normala" färgseendet, medför även individuella variationer inom de grupper som diagnosticeras som "anomala". Pokorny och Smith (1977) konstaterar till och med att "interobserver variability is characteristic of individuals having anomalous color vision". Det är exempelvis endast i statistisk mening som dikromater "alltid" accepterar trikromaters färgmatcher, påpekar de.
Ytterligare en variation i det normala färgseendet är den som man på senare tid funnit hos mödrar till anomala söner. Dessa kvinnor, som på den ena av sina två X-kromosomer bär ett anlag för färgseendeanomali, har traditionellt räknats som "normalseende", eftersom de klarar sedvanliga färgsinnestest. Nagy och hans medarbetare (1981) har emellertid funnit intressanta avvikelser från det normala hos dessa s.k. "heterozygota" kvinnor. En grundprincip i den trikromatiska modell för färgseende, som jag inledningvis skisserade, är additivitet. Det innebär att om jag har funnit en match mellan två fysikaliskt olika stimuli och så adderar ett godtyckligt ljus till båda, så bibehålls matchen. Exempelvis vid en Rayleigh-match (y=r+g); om på båda fälten superponeras ett blått ljus, så ser de fortfarande lika ut. Det visade sig vara fallet för alla män i den undersökning av additivitet som Nagy et al. gjorde, men däremot inte för 4 av de 21 kvinnliga försökspersonerna. Dessa visade sig vid förfrågan mycket riktigt ha färgblindhet i släkten. Att additivitet inte gäller för dessa heterozygota kvinnor skulle kunna förklaras om man antar att de har fler än de tre sedvanliga pigmenten i sina receptorer. Förslagsvis de tre normala plus det anomala pigment som motsvarar den anomala gen de bär. Dessa rön pekar med andra ord på den intressanta möjligheten att det skulle kunna förekomma personer med ett avvikande färgseende, som bestode i att de hade flera skilda slag av receptorer än normalt, dvs att de vore tetrakromater.
TEMPORALA FENOMENEtt välkänt och förbryllande fenomen är de färger som kan varseblis i samband med roterande svart-vita mönster. På den s.k. Benham-snurran omväxlar sektorer av svart, vitt och ränder. Den ordning i vilken dessa tre element avlöser varandra, när skivan roterar, avgör om en observatör ser ränderna skimra i mörkblått, grönt, gult eller rött. Eftersom dessa kulörer uppträder till följd av en periodiskt växlande stimulering - och inte har att göra med fotoreceptorernas absorptionsspektra - kunde man tänka sig att färgblinda (dikromater) inför Benhamsnurran skulle uppleva rött och grönt, även om de aldrig gjort det vid sedvanliga optiska stimuli.. C.W.White, G.R.Lockhead och N.J.Evans (1977) undersökte denna möjlighet, med en metod liknande den tidigare beskriva av Shepard och Cooper, dvs genom skattning av grad av likhet mellan parvis iakttagna kulörer. Även om en färgblind försöksperson faktiskt spontant utbrast: "That line is definitely coloured, but I don't know what to call it" och "Those lines are greener than grass", så var resultatet som helhet nedslående. De färgblinda var inte kapabla att göra någon åtskillnad mellan vad en normalseende såg som rött och grönt. Det verkar som om de uppfattar "Benham-färgerna" på samma sätt som andra färger. Detta faktum ger stöd för tesen att den färganomales defekt inte bara är en fråga om frånvaron av ett slag av fotopigment, utan även manifesterar sig på nevronal nivå.
Välj i ÖVERSIKTEN eller gå direkt till NÄSTA KAPITEL AFS5.htm |